I investigate the cognitive mechanisms supporting adaptive control over behavior. To be adaptive over both evolutionary timescales and more momentary ones, and over both health and disease, these cognitive control mechanisms must balance constraints common to many resource-limited systems. I take inspiration from such systems, using formal models to refine hypotheses about how cognitive control may address these fundamental constraints. I then test these hypotheses with a convergent cognitive neuroscience approach. I test children and adults using physiological techniques like pupillometry and EEG, and cutting-edge technologies like fMRI and TMS, all building on a computational foundation in neural networks and Bayesian modeling. In my former role in Pfizer's Worldwide R&D department, as well as my current role at Roche's Basel Innovation Center, I use knowledge from my research to assess the impact of novel compounds on cognition and the neural circuits supporting it.
Christopher H. Chatham
Computational Cognitive Neuroscience of Adaptive Control
<
>
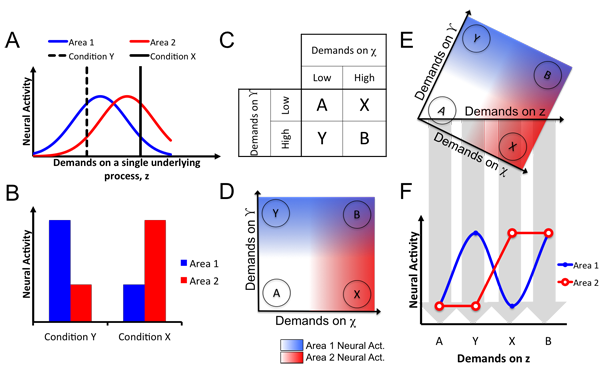
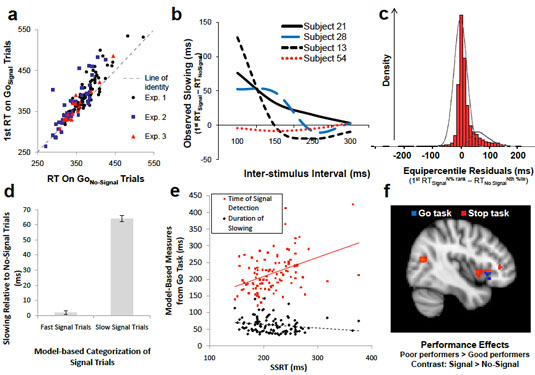
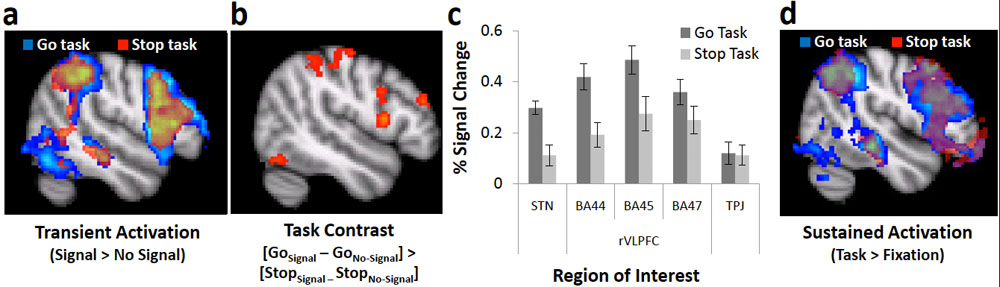
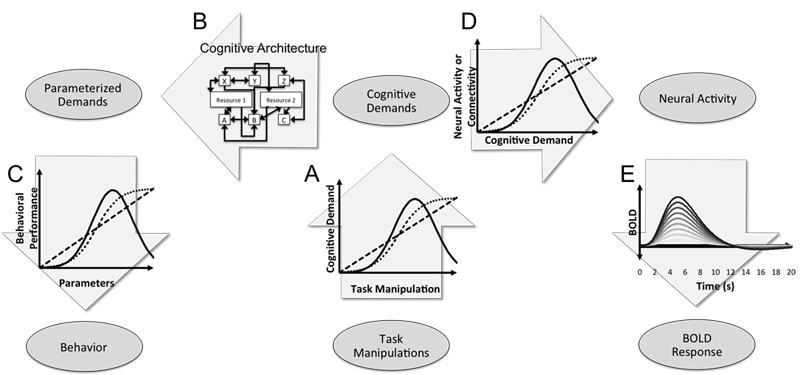
In a forthcoming book chapter on "How to Test Cognitive Theory With fMRI", we describe clear inferential risks that confront even the most ideal experimental outcomes - such as the cross-over double dissociation shown above. Inferential ambiguities like the ones depicted here motivate our many recommendations for very specific experimental design in cognitive neuroimaging. See my publications for more.
One aspect of my work focuses on how we stop unwanted behaviors. I hybridized a few statistical models of behavior - namely, race models and mixture models - to reveal new subcomponents to measures of this ability. Our work seems to imply that measures of stopping tend to require control for a very different reason than many scientists (currently) think. See my publications for more.
Do influential response inhibition tasks actually assess anything intrinsically related to stopping? Some of our work strongly challenges this notion. See my publications for more.
This in-progress neural network simulation is already showing patterns of performance that match humans in tests of selection from working memory. In particular, the model naturally shows an increasing dispersion of settling times that also characterize the response time distributions of humans, when they are asked to select a subset of information currently in mind. Stay tuned for more.
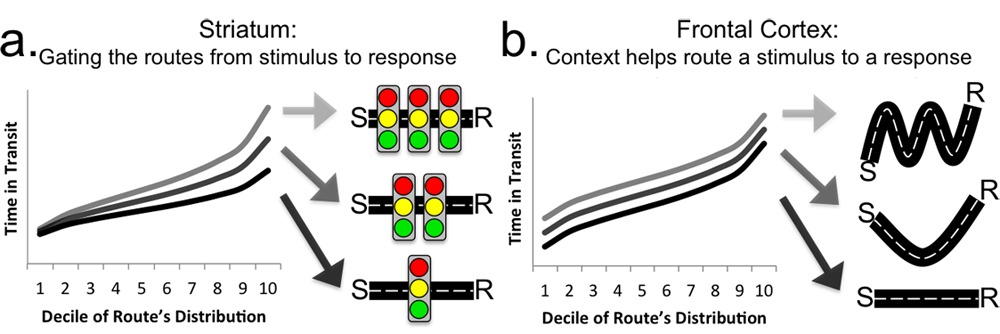
Our understanding of the neural basis of cognition can be informed by the study of other complex systems. One particularly unexplored parallel is with the study of traffic (and other queued networks). In work currently under-review, we have used traffic as an intuitive way of describing the dynamical contributions of frontal and striatal systems to working memory performance.
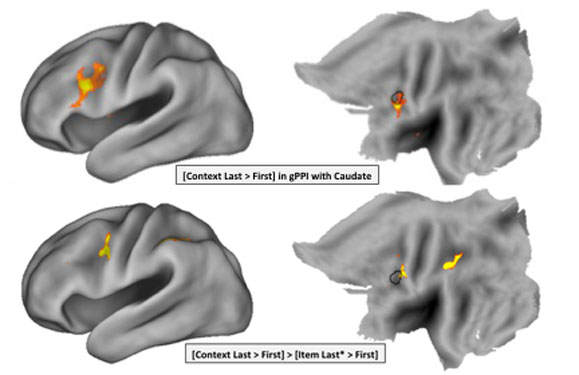
We can keep multiple items actively "in mind" concurrently. This would pose a problem if all such items were simultaneously competing for attention and influence over action. One potential solution to this problem of concurrency is to grant certain items a priviledged influence over attention and behavior. We hypothesized that this ability might rely on a particular corticostriatal circuit (specifically, the dorsal pre-PMd and caudate). In work currently under review, we report evidence for the neural network models that drove this prediction.
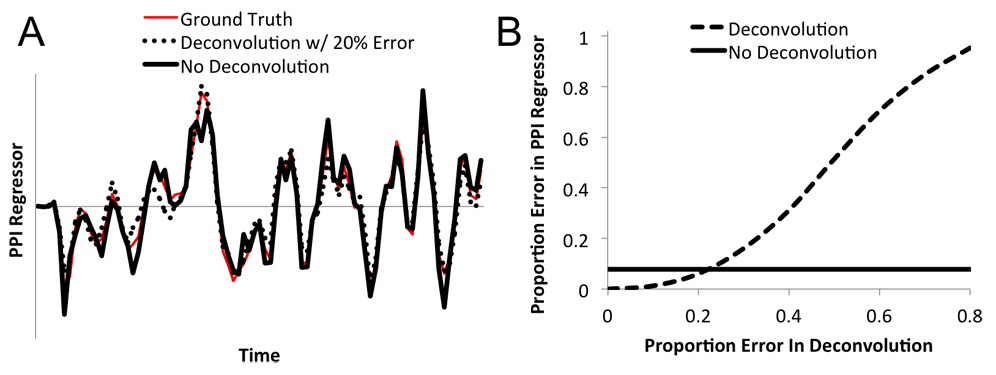
One of the tools I use is the psychophysiological interaction (PPI) - a neuroimaging technique that can reveal task-dependent changes in connectivity. Major software packages accomplish this procedure in different ways, with some degree of controversy. Surprisingly this difference had not been directly assessed. We used Monte Carlo techniques to show the risks and potential of each approach, under simplifying but plausible assumptions. See my publications for more.
This figure makes explicit several inferential steps in cognitive neuroimaging (right side) as they compare to those in classical cognitive studies (left side). See my publications for more.
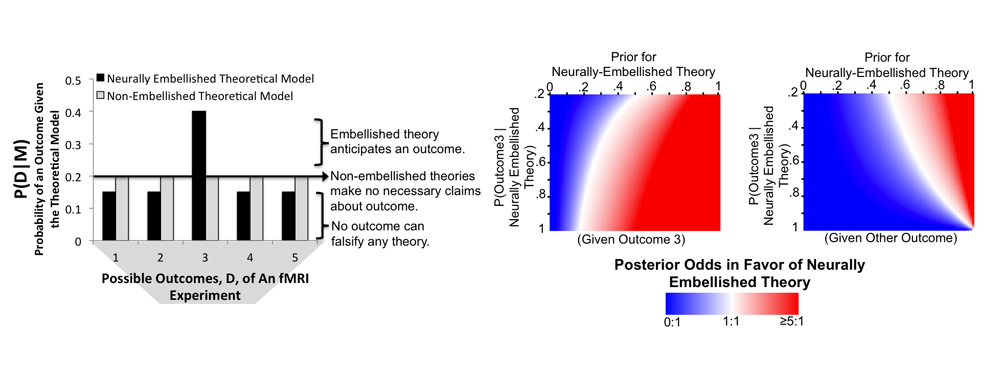
It is important to seriously confront methodological criticisms of cognitive neuroscience, and several particularly outspoken critics have noted something they term the "consistency fallacy." This so-called fallacy occurs when neuroimaging results consistent with a particular cognitive theory are taken as evidence for it, even if no outcome could possibly falsify the theory. The figure above is adapted from our argument that there is actually no such fallacy, at least from a Bayesian conception of evidence.
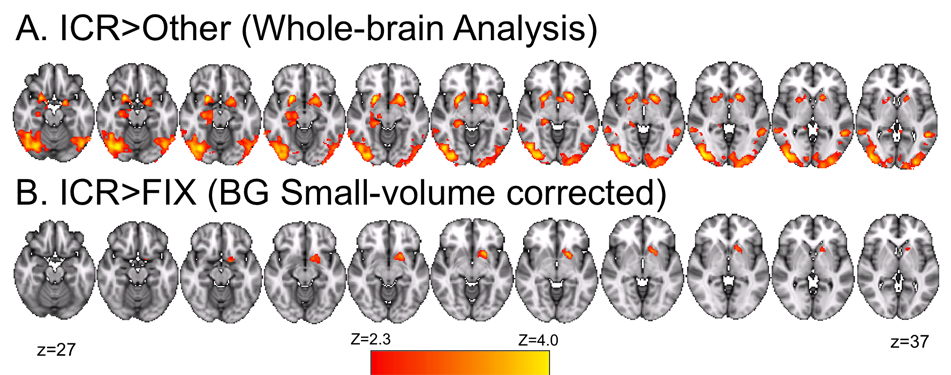
In model-based fMRI, one lets loose a biologically-plausible algorithm on some task that humans are asked to learn in an MRI scanner. Parameters in the algorithm that vary along with hemodynamic activity in the brain can provide hints about neurocognitive function. Above I show parts of the brain that seem to "veto" instructions to reallocate working memory capacity, in a way that corresponds with simple reinforcement learning principles. See my publications for more details.
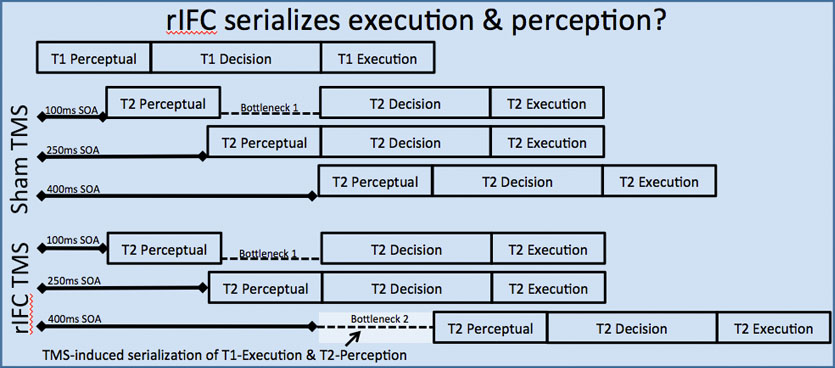
Several years ago, a very interesting paper using transcranial magnetic stimulation was published in the Proceedings of the National Academy of Sciences. Upon careful inspection, it seems to show a pattern of human performance that has not been observed in 50+ years of research. In the future I hope to replicate this pattern and explore the implications for our understanding of adaptive cognitive control. See my publications for a commentary on the study and fuller description of its fascinating findings.
A central question in cognitive psychology concerns the format of mental representations. One can address this question by assessing the similarity of distributed patterns of neural activity across stimuli that varying in quantifiable ways. An advantage to this "representational similarity" approach is that it can be conducted on realistic computational models as well as real people. Above, I show the representational similarity of various items in the "working memory" of a neural network model of the corticostriatal system. This model was the first to be capable of performing the challenging n-back working memory task. See my publications for more.
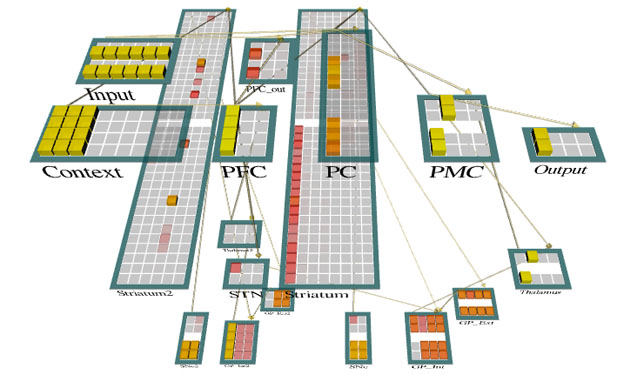
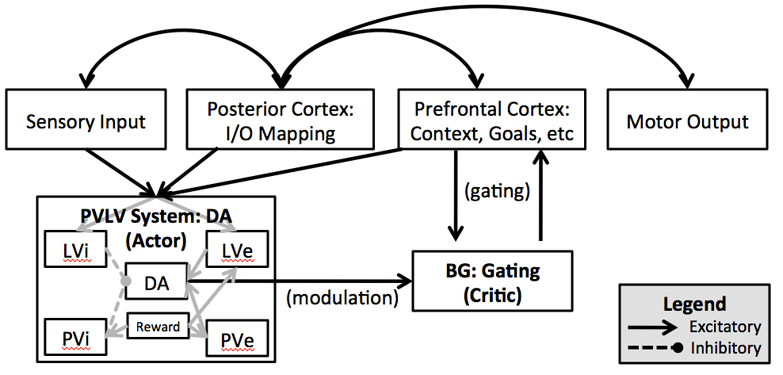
This schematic depicts an adaptation of the prefrontal basal ganglia working memory architecture (pioneered by O'Reilly, Frank and colleagues) used to simulate a challenging test of cognitive control over working memory: the n-back. See my publications for more.
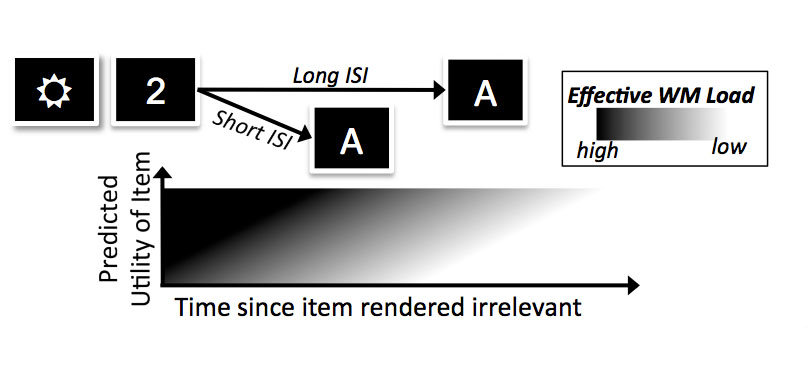
The storage space in working memory is highly capacity-limited; adaptive control over its contents is therefore key, and resources should be reallocated from no longer relevant information. This schematic illustrates a model we developed to assess whether the timecourse of working memory reallocation is associated with the processing of information utility. See my publications for more.
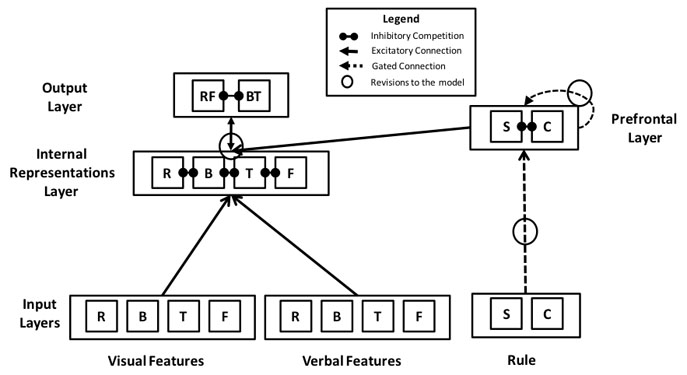
Schematic of the neural network used to simulate the role of working memory updating in the surprising behavior of preschoolers - who spontaneously adopt an "opposites game" rather than simply switch from sorting cards by color to sorting them by shape See my publications for an exploration of these issues.
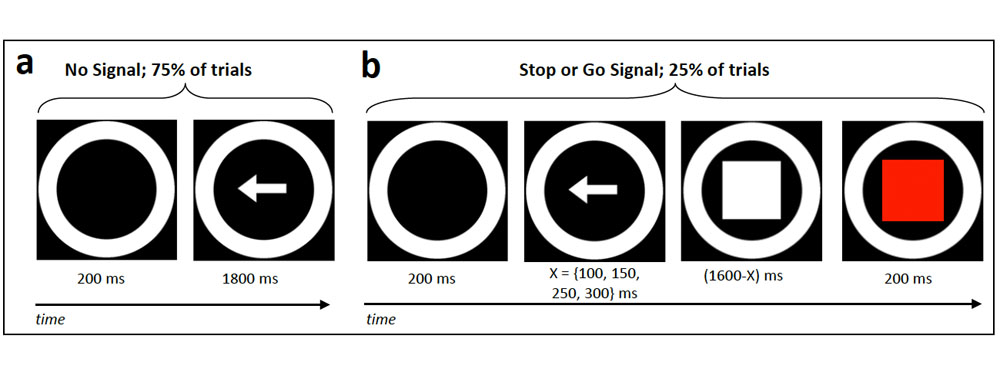
In the Stop Signal task, the majority of trials involve pressing one of two buttons in response to one of two arrows - quite simple. But a small minority of trials involve an unpredictable and sudden signal to stop all behavior. Many have discussed results from this task as though they reflect this demand to stop behavior, but our work - which simply compares the Stop Signal to another infrequent "Go Signal" demanding no stopping at all - shows that this is very unlikely to be the case. See my publications for more.
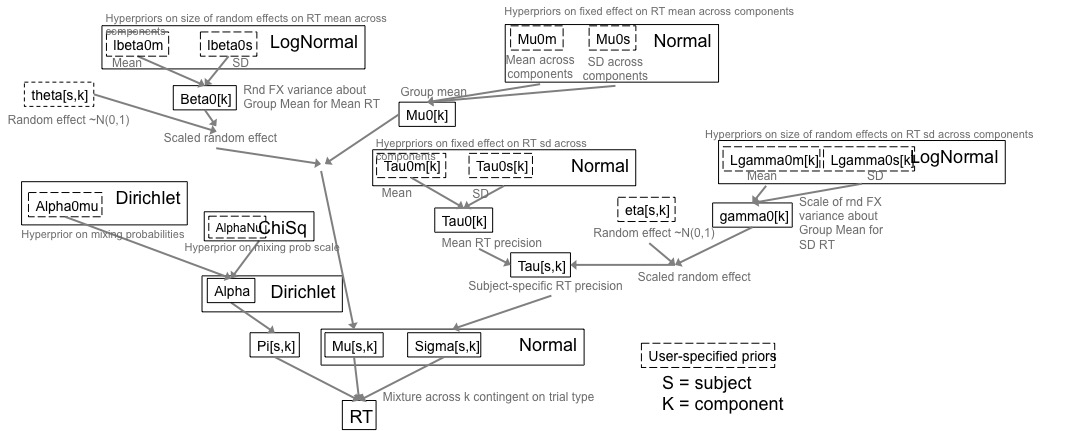
I recently implemented this hierarchical Bayesian mixture model - a class of models that are typically difficulty to evaluate due to their propensity to show poor mixing. I adapted the model from that of Almond et al., 2009, for use in a new manuscript examining the intersection of proactive vs. reactive control strategies and input vs. output gating mechanisms for updating working memory.
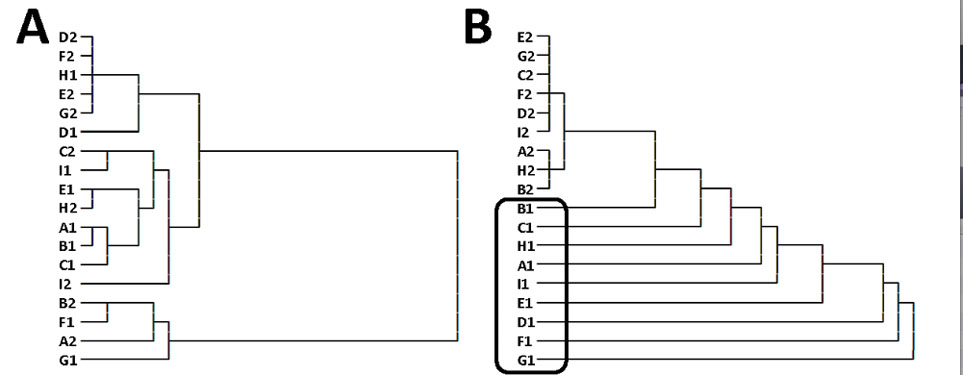
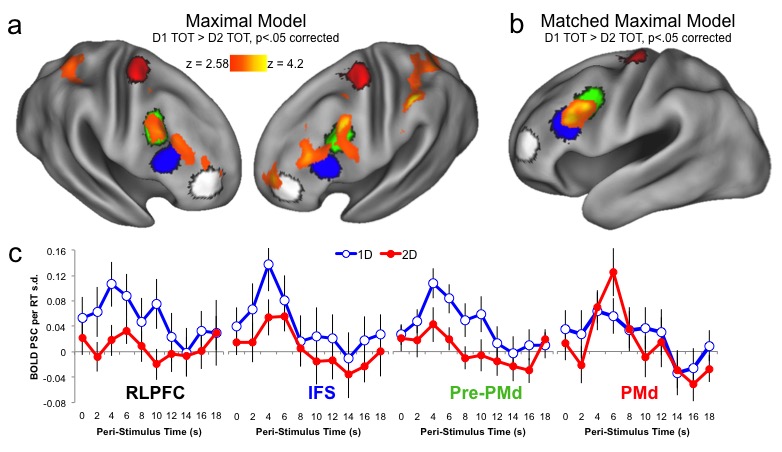
An often overlooked inferential risk in cognitive neuroscience is that a statistical pattern know as the "double dissociation" is not, as commonly presumed, a guarantee of dissociable cognitive functions. Sophisticated recent methods for overcoming this limitation (e.g., state-trace methods) have likewise come under fire. We've made some new inroads to the problem - this is a glimpse of a demonstration of our approach on data from a hierarchical cognitive control task. We hope to fully report our approach and its potential in a (currently under-review) publication.